
BRAU- und MESSTECHNIK
Brausystem, eingesetzte Messverfahren und weitere Techniken im Überblick
BRAU- und MESSTECHNIK
Brausystem, eingesetzte Messverfahren und weitere Techniken im Überblick
BIOSCIENCE- BIOLOGIE der GÄRUNG
ENERGIESTOFFWECHSEL, GÄRUNGEN - Energiegewinn ohne Sauerstoff
SAUERSTOFF - Feind oder Freund des Brauers?

Inhaltsverzeichnis

Abb. 1. Gärungen als Notmechanismus zur Energiegewinnung. [Quelle: Weber, Biologie Oberstufe. 2. Aufl. (2009), S. 108] 05.08.2021
1. ENERGIE: ohne Energiestoffwechsel kein Leben TOPINFO wichtige BEGRIFFE: siehe GLOSSAR Biochemie
Brennstoffe bzw. Energieträger sind uns im Alltag bestens vertraut: Benzin zum Autos fahren, Elektrizität für Eisenbahnen, Industrie und alle Arbeitsleistungen zuhause wie Kochen, Staubsaugen, Lichterzeugung, Warmwassererzeugung, Betrieb elektronischer Geräte wie Computer, Kamera, Handy/Smartphone, Holz und Kohle für Heizzwecke.
Auch Lebewesen vom Einzeller (Bakterien, Hefen) bis zum Vielzeller (Pflanze, Tier, Mensch) brauchen ständig Energieträger, entweder energiereiche Nahrungsmittel oder - exklusiv nur Pflanzen und gewisse Bakterienarten - Sonnenlicht für die Fotosynthese. Die Nahrungsmittel werden primär als Betriebsstoffe für den Energienachschub (Mensch: ca. 80-90%) im Stoffwechsel umgewandelt (Energiestoffwechsel --> universelle biologische Energiewährung ATP [Adenosintriphosphat]). Nur einer kleiner Anteil wird als Baustofflieferant für die Grundbausteine aller Körpersubstanzen eingesetzt (Baustoffwechsel --> Aminosäuren der Eiweisse, Fettsäuren, Kohlenhydrate, cf. Kurzinfo). Dieser ständig notwendige Energienachschub wird für alle biologischen Arbeiten benötigt, z.B. Erhaltung der Körperwärme, Aufrechterhaltung körperlicher Funktionen (z.B. Gehen, Muskeltätigkeit, Verdauung, Herzschlag, Organfunktionen etc.), Aufrechterhaltung geistiger Funktionen, Wachstum, Fortpflanzung, Regeneration verlorengegangener Körperbestandteile (z.B. Haut, Haare, Fingernägel), Stoffwechseltätigkeiten wie Aufbau der Zellen/Organe. Auch einzellige Lebewesen benötigen für manch ähnliche Funktionen laufend Energie.
Energie kann grundsätzlich auf 4 Arten gewonnen werden, nämlich durch
-
Fotosynthese - Aufbau von Energieträgern wie Zucker/Stärke C6H12O6/(C6H12O6)n mittels Sonnenenergie aus Wasser H2O und Kohlenstoffdioxid CO2 - nur Pflanzen und wenige Bakterienarten (z.B. Cyanobakterien, allg. phototrophe Bakterien) = sog. Produzenten: 6 CO2 + 6 H2O + Sonnenenergie --> C6H12O6 + 6 O2 (Kurzinfo, cf. auch Glossar).
-
Aerobe Atmung - klassische Atmung mit Luftsauerstoff O2, z.B. Abbau von Zucker/Stärke zu Wasser und Kohlenstoffdioxid: C6H12O6 + 6 O2 + 32 (ADP + Pi) --> 6 CO2 + 6 H2O + 32 ATP) - Atmer, Aerobier: alle Tiere, Mensch, Pflanze, Bakterien, Hefen (Kurzinfo; cf. auch Glossar)
-
Anaerobe Atmung - Atmung ohne Sauerstoff - nur gewisse Bakterien, z.B. Methanbakterien (Kurzinfo).
-
Gärungen - Atmung ohne Sauerstoff durch unvollständigen Abbau eines organischen Moleküls, z.B. alkoholische Gärung durch Zuckerabbau: Zucker C6H12O6 + 2 (ADP + Pi) --> 2C2H5OH + 2CO2 + 2 ATP - Gärer, Anaerobier: Mikroorganismen wie Bakterien und Hefen (Kurzinfo).
2. GÄRUNG: Energiegewinnung ohne Sauerstoff
Bei Mikroorganismen wie Bakterien und Hefen kommen verschiedene Formen der Gärung vor. Zwei bekannte und wichtige Formen sind die alkoholische Gärung (Ethanolbildung, überwiegend durch Hefepilze und Bakterien) und die Milchsäuregärung (hauptsächlich durch Milchsäure-bakterien). Daneben gibt es noch 6 weitere wichtigere Gärungsformen: 2,3-Butandiol-Gärung, Ameisensäuregärung, Buttersäuregärung, Gemischte Säuregärung, Homoacetatgärung und die Propionsäuregärung. Allen Gärungen ist gemeinsam, dass sie vom Endprodukte der Glykolyse, der Brenztraubensäure (= Pyruvat) ausgehen und nur unter anaeroben Bedingungen stattfinden, d.h. sie sind nicht auf Sauerstoff angewiesen, liefern aber bedeutend weniger Energie in Form von "biologischem Treibstoff", den ATP-Molekülen.
Mikro-
organismen
Infoeinschub
Mikroorganismen (Mikroben, Kleinstlebewesen) sind mikroskopisch kleine Organismen, die als Einzellebewesen nicht mehr von blossem Auge sichtbar sind. Sie kommen meist als Einzeller vor, es gibt aber auch wenigzellige Mikroorganismen. Zu den Mikroorganismen zählen Bakterien (z. B. Milchsäurebakterien), Archaeen (z.B. Methanbildner), Mikropilze (z. B. Backhefe, Bierhefe), Mikroalgen (z. B. Kieselalgen, Grünalgen) sowie tierische Einzeller (Protozoen, z. B. das Pantoffeltierchen, Amöben oder der Malaria-Erreger Plasmodium). Viren sind ohne eigenen Stoffwechsel und daher keine Lebewesen, werden aber trotzdem als ein Teilgebiet der Mikrobiologie angesehen (Virologie), ebenso wie die Prionen (infektiöse Eiweisspartikel) (Kurzinfo Mikroorganismen).
Bakterien und Archaeen (= Prokaryoten) sowie Viren sind kernlos, Mikropilze, Mikroalgen und Protozoen sind kernhaltig (= Eukaryoten).
Mikroorganismen nehmen im Stoffkreislauf der Natur wichtige Funktionen wahr: Zum einen bilden sie als Produzenten (z. B. Mikroalgen) die Grundlage vieler Nahrungsketten, zum anderen bauen sie als Zersetzer (Destruenten) organische Materie zu anorganischen Stoffen ab. Einige Mikroorganismen sind für die Ernährung von Bedeutung, einige für erwünschte Stoffumwandlungen (beispielsweise Antibiotika-Produzenten), andere sind Parasiten und Erreger von Infektionskrankheiten. In der Biotechnologie gewinnen die Mikroorganismen ständig an Bedeutung. Die Mehrzahl der Mikroorganismen ist für uns Menschen nützlich bis harmlos, nur wenige sind für uns heikel (Infektionserreger).
Mikroorganismen übertreffen alle anderen Lebewesen bei weitem an Zahl und stellen mit 70 Prozent den grössten Anteil an lebender Materie (Biomasse).
Stoffwechsel der Hefen: Hefen können sowohl als Atmer (Aerobier) wie auch als Gärer (Anaerobier) leben, sie sind somit fakultative Aerobier. In Gegenwart von Sauerstoff und sehr geringen Zuckerkonzentrationen veratmen die Hefen Zucker vollständig zu CO2 und Wasser und gewinnen dabei genügend biologische Energie, die sie zum Wachstum und zum Aufbau neuer Zellen nutzen (Zellvermehrung). Ist die Luftzufuhr (Luftsauerstoff) für Hefen vorhanden und herrscht ein Glukoseüberschuss vor, der die Aufnahmekapazität der Atmungskette überschreitet (Konzentrationen von mehr als 0.1 g/L), so schalten sie ihren Stoffwechsel auf die alkoholische Gärung um (= Crabtree-Effekt, benannt nach dem Biochemiker H.G. Crabtree). Ist kein Sauerstoff vorhanden, aber Zucker, so vergärt sie den Zucker zu Ethanol und CO2. Diese anaerobe alkoholische Gärung hilft ihnen, eigentlich lebensfeindliche Umweltbedingungen zu überleben, obwohl das energetisch höchst ungünstig ist (cf. unten). Der französische Mikrobiologe Louis Pasteur (1822-1895) entdeckte 1861, dass Hefe ohne Sauerstoff mehr Glukose verbraucht als in O2-reicher Umgebung: das ist der sog. Pasteur-Effekt. Die Hefen müssen unter anaeroben Bedingungen weit mehr Zuckermoleküle verarbeiten als unter aeroben Bedingungen, um die Energieverluste zu kompensieren. Fazit: Ob die Hefe atmet oder gärt, hängt also primär von der Zucker- und Sauerstoffversorgung ab. Im Brauprozess wird durch den hohen Zuckergehalt der Bierwürze sofort der Crabtree-Effekt wirksam, d.h sie schaltet auf die anaerobe alkoholische Gärung um.
Milchsäuregärung und Alkoholische Gärung (Fermentationen: M-Gärung, A-Gärung)
Beide Gärungsformen sind in der Bierherstellung von grosser Bedeutung. Die alkoholische Gärtung kommt bei jedem Biertypus zum Einsatz, während die Milchsäuregärung gezielt nur bei den "Sauerbieren" (engl. sour beer) wie Berliner Weisse, belgische Lambic oder Geuze-Biere eingesetzt wird (Info, Zusatzinfo).
Beim effizientesten, also ertragsreichsten Abbau energiereicher Nährstoffe wie Stärke via Zuckereinheiten (z.B. Glukose) durch die aerobe Atmung (syn. Respiration, Veratmung, Atmungsstoffwechsel, aerobe Atmungskette) mit Luftsauerstoff werden pro Glukosemolekül 32 biologische Energiewährungs-einheiten in Form des ATP-Moleküls gewonnen: C6H12O6 + 6 O2 + (32 ADP, 32 Pi) --> 6 CO2 + 6 H2O + 32 ATP. Steht aber kein oder nicht ausreichend Sauerstoff zur Verfügung, muss die Zelle ihren "Abbaustoffwechsel" (Katabolismus) ändern. Einen Ausweg stellen die Gärungen dar - die abbauenden Prozesse verlaufen zwar nicht vollständig zum CO2 und liefern nur relativ wenig ATP, ermöglichen es der Zelle aber, unter sauerstofffreien (aeroben) Bedingungen weiterzuleben: Anerobe alkoholische Gärung C6H12O6 + (2 ADP, 2 Pi, 2 H+) --> 2 Ethanol C2H5OH + 2 CO2 + 2 ATP + 2 H2O. Die Energieausbeute von nur 1/16 im Vergleich zur aeroben Atmung (2 ATP vs. 32 ATP) zwingt die Gärerzellen wie Hefen, wesentlich rascher Stärke bzw. Zucker zu verstoffwechseln, um die gleichen biologischen Funktionen erfüllen zu können wie die Aerobier z.B. Zellteilung/ Zellvermehrung und Aufbau neuer Zellen. Dabei wird in Kauf genommen, dass sogar energiereiche Produkte aus der Zelle ausgeschieden und nicht weiter genutzt werden können wie eben Ethanol! Dass Ethanol immer noch energiereich ist, wissen wir beim Anzünden von Brennsprit z.B. beim Fondue. Dieser für Mikroorganismen sehr ineffiziente Stoffwechselvorgang zur Energiegewinnung ist für den Menschen von grossem Vorteil, nämlich die Ausscheidung der für die Mikroorganismen "unnützen" Endprodukte wie Alkohol oder Milchsäure in grossen Mengen. Diese Tatsache wird besonders in der Biotechnologie bei manchen mikrobiellen Produkten zu Nutze gemacht.
Ein genauerer Blick auf die M- und A-Gärung
Beide Formen der Gärung beginnen mit dem biochemischen Stoffwechselprozess der Glykolyse ("Zuckerabbau"). Dabei werden pro Glucosemolekül (Traubenzucker, C6H12O6) zwei Moleküle des biologischen Energieträgers ATP gebildet sowie Pyruvat (auch Brenztraubensäure genannt, ein C3-Körper H3C-CO-CO-OH, Summenformel C3H4O3, cf. Abb. alkoholische Gärung).

siehe auch: Mikrobenzirkus

Abb. 2. Wege der Energiegewinnung.
Zuckerverstoffwechslung läuft zunächst über die Glycolyse ("Zuckerauflösung") von C6 (Glucose) zum C3 (Pyruvat).
Mit Sauerstoff O2 kann die Zellatmung einsetzen (--> liefert viel Energie als ATP). Ohne O2 kann die Gärung ablaufen, die wenig ATP liefert, z.B. in Hefen als alkoholische Gärung (Endprodukte C2 [Ethanol] und CO2), oder in Bakterien als Milchsäuregärung (C3 [Lactat bzw. Milchsäure]).
GLYKOLYSE Zuckerabbau Infoeinschub Info

Abb. 3. Ein ganz entscheidender Stoffwechselpfad: Die Glykolyse.
Sowohl die aerobe Atmung wie auch die Gärung beginnt mit der Zuckerzerlegung, der Glykolyse.
Der Glukoseabbau verläuft über 9 Zwischenprodukte, die alle durch insgesamt 10 Enzyme (Nr. 1-10) ermöglicht und beschleunigt werden.
Die Glykolyse hört mit einer C3-Substanz, dem Pyruvat (Brenztrauben-säure) auf. Diese Substanz wird sowohl in der anschliessenden aeroben Atmung als auch in der anaeroben Gärung (Fermentation) weiter verstoffwechselt.
Die anschliessende alkoholische Gärung wie auch die Milchsäuregärung benötigen weitere Enzyme (Nr. 11 bzw. Nr. 12-13)
[Quelle: Brock Mikrobiologie kompakt, 13. Aufl. (2015), S. 96]
Bilanz des Zuckerabbaus (Glykolyse): C6H12O6 [Traubenzucker = Glukose] + 2 ADP [entwerteter biol. Energieträger]+ 2Pi [Phosphat] + 2 NAD+ [Wasserstoffüberträger = Elektronenüberträger] --> C3H4O3 [Pyruvat, wichtiger Zwischenstoff als "Drehscheibe" im Gesamtstoffwechsel] + 2 ATP + 2 NADH + 2 H+ (abgekürzt NADH2) + 2 H2O.
Vereinfacht: C6H12O6 --> 2 C3H4O3 + 2 ATP
Nach der Glykolyse wird ein Mechanismus benötigt, der die Regeneration des Wasserstoff- bzw. Elektronenüberträgers NAD+ aus NADH steuert. Ohne diese Regeneration wäre der Zellvorrat an NAD+ schnell erschöpft und die Gärung käme zum Erliegen. Im Unterschied zur aeroben Zellatmung, bei der das in der Glykolyse entstehende NADH seine Elektronen auf die sog. Atmungskette zur sehr effizienten Bildung von ATP überträgt, werden sie bei der Gärung an das Zwischenprodukt Pyruvat C3H3O3- abgegeben.
Bei der alkoholischen Gärung wird Pyruvat C3 in zwei Reaktionsschritten zum Alkohol Ethanol C2H5OH umgewandelt. Zunächst wird aus dem C3-Molekül Pyruvat ein C als CO2 abgespalten, es entsteht das C2-Molekül Acetaldehyd C2H4O, das im zweiten Reaktionsschritt durch NADH (Elektronen- bzw. H-Überträger) zu Ethanol reduziert wird. Dieser Vorgang wird zum Bierbrauen, zur Wein-/Schaumweinherstellung, für Cidre, Sake, Branntweine (Rohstoffbasis Wein --> Weinbrand, Cognac, Armagnac; Zuckerrohrsaft oder Melasse --> Rum, Arrak); Getreide --> Korn, Whisky); Kartoffeln --> Wodka; Obst --> Obstwasser, Obstgeist; Agavenpulpe --> Tequila u.a.), Reiswein (Sake) und über das abgespaltene Gas CO2 auch zum Backen genutzt.
Abb. 4. Gärungen beginnen als Zuckerabbau (C-Atome: schwarze Kugeln; H- bzw. e-
[Elektronen]-Überträger] NAD+ gelb. Oben: Zuckerabbau, unten: 2 wichtige Gärungen.
Links: Milchsäuregärung. Bilanz: Glucose + 2 (ADP + Pi) --> 2 Lactat + 2 ATP
Rechts: Alkoholische Gärung. Bilanz: Glukose + 2 (ADP + Pi) --> 2 Ethanol + 2CO2 + 2 ATP
[Quelle: Markl Biologie. 1. Aufl. (2010), S. 113 (modifiziert). Abb. mit mehr Details: hier klicken!]

Milchsäuregärung: Die Milchsäuregärung wird nicht nur von Mikroorganismen bei Sauerstoffabwesenheit angewandt, sondern auch bei den meisten Tieren und beim Menschen (!).
Beispiel: Sie erklimmen mit dem Mountainbike einen steilen Berg, der kleinste Gang ist bald nicht mehr ausreichend. Sie müssen absteigen, aber der Berg ist noch nicht überwunden. Ihr Puls und ihre Atemfrequenz sind bald am Limit. Ihre Oberschenkel beginnen nun auch noch zu brennen: Das tut richtig weh! Jetzt bleibt nichts mehr übrig als abzusteigen und auszuruhen. Was ist da physiologisch betrachtet passiert? Die Muskelzellen ihrer Beine mussten von aerober Energiegewinnung (klassische Atmung, siehe oben) auf anaerobe Energiegewinnung umschalten, da nicht mehr ausreichend Sauerstoff O2 via Blutbahn geliefert werden konnte. Aber das ist nur eine kurzfristige Notlösung, denn in Abwesenheit von Luft-O2 kommt die Reaktionskette der aeroben Atmung, die sog. Atmungskette der Zellatmung fast vollständig zum Erliegen.
In der Milchsäuregärung nun wird das Endprodukt der Glykolyse, das Pyruvat, zu Milchsäure (Lactat) reduziert und gleichzeitig NADH zu NAD+ oxidiert (vgl. Abb. 4). Physiologisch betrachtet ist aber die Bildung von Milchsäure eine Sackgase, denn diese Säure muss aus den Muskeln über das Blut zur Leber gebracht werden. Der Lactatspiegel wäre unmittelbar nach dem Absteigen vom Mountainbike deutlich erhöht gewesen. Die dadurch angehäufte "Sauerstoffschuld" muss in der Erholungsphase wieder abgetragen werden: d.h. das energiereiche Lactat H3C-HCOH-COO- (Summenformel C3H5O3-) wird in der Leber wieder in Glukose zurückverwandelt. Doch das ist nicht nur zeitraubend, sondern kostet auch noch Energie in Form von ATP und dafür wird viel Sauerstoff gebraucht: man atmet also noch lange heftig.
Mikroorganismen wie die Milchsäurebakterien scheiden aber einfach die "störende" Milchsäure ins Aussenmedium aus: die Milch wird sauer und bricht dann (Eiweisse werden durch Säuren ausgefällt), die Bierwürze wird sauer, bis ab einem bestimmten tiefen pH-Wert (Säuregrad) auch die Mikroorganismen in ihrer Stoffwechseltätigkeit blockiert werden, da die verantwortlichen Enzyme auf den pH-Wert empfindlich reagieren. Enzyme haben immer ein pH-Optimum, z.B. die Stärke abbauenden Amylasen um pH 5.2- 5.7.
Beteiligte Organismen: Milchsäurebakterien wie Lactobacillus acidophilus, L. casei, L. lactis, L. bulgaricus, L. helveticus, L. delbrueckii, L. bifidus, L. brevis, Leuconostoc mesenteroides, Streptococcus pyogenes, u.a.
Die Milchsäuerung wird genutzt: 1. Milchsäuerung (Sauermilch, Sauerrahm, Joghurt, Kefir, Buttermilch), 2. Gemüse (Sauerkraut), 3. Futtermittel (Silage).
ATP NADH/H+ Schlüsselmoleküle: Energieträger Infoeinschub
ATP ist ein Schlüsselmolekül in der direkten Energielieferung, die universelle "biologische Energiewährung" schlechthin. ATP ist der zentrale und entscheidende Energieträger aller Organismen! Immer wenn es darum geht, Energie kurzfristig zu transportieren, sher kurzfristig zu speichern oder andere Moleküle in einer Stoffwechselkette (= hintereinander ablaufende biochemische Reaktionen wie z.B. Glykolyse) zu aktivieren, also "reaktionsfähig" zu machen, nutzen alle Zellen fast immer Adenosintriphosphat ATP. Das Molekül besteht aus einem Baustein Adenosin (= eine Aminosäure Adenin, Baustein von Eiweissen und dem Zucker Ribose) und einer 3er-Kette von Phosphatgruppen. Die Besonderheit des ATP liegt in der 3. Bindung der Phosphatgruppen: diese kann durch Anlagerung von Wasser leicht gespalten werden (= hydrolytische Spaltung). ATP ist folglich relativ instabil und die viele bei der Hydrolyse freiwerdende Energie kann für andere Reaktionen in Form einer energetischen Kopplung (vgl. Glossar Biochemie > energetische Kopplung) genutzt werden: ATP + H2O <--> ADP + Pi + 30.5 kJ/mol. ATP gibt auch bereitwillig eine Phosphatgruppe an andere Moleküle ab und hat somit ein hohes Gruppenübertragungspotenzial für Phosphat (= Phosphorylierung; vgl. Glossar Biochemie > Glykolyse, Abb.).
Die Bedeutung des ATP für den Menschen zeigt sich auch
quantitativ: Ein erwachsener Mensch produziert und hydrolysiert
jeden Tag etwa 40 kg ATP. Bei einer Gesamtmenge von nur ca. 50 g
ATP pro Mensch entspricht das einer hohen Regeneration:
durchschnittlich in jeder Minute einmal, also mehr als 800x pro
Tag. Dieser enorme Umsatz wird zu 90% von den "Atmungszentren"
der Zelle, den Mitochondrien bereitgestellt. Diese Zellorganellen
nehmen das ADP und das Phosphat Pi auf, das bei der Hydrolyse
von ATP anfällt und regenerieren daraus ATP. Dies ist aber nur bei
der aeroben Zellatmung möglich, wenn pro 1 Glukosemolekül 32
ATP-Moleküle anfallen. Bei der sauerstofffreien Notlösung zur
Energiegewinnung, der alkoholischen Gärung, fallen nur gerade 2
ATP-Moleküle pro Glukose an. Kein Wunder, dass die Hefezellen
unter Luftabschluss wie verrückt den Zucker umsetzen müssen,
um genügend ATP für ihr Überleben gewinnen zu können!
Abb. 5. Bau der biologischen Energiewährung ATP, seine Spaltung unter Wassereinfluss (Hydrolyse) in ADP und Pi.
[Quelle: Markl Biologie, 1. Aufl. (2010), S. 67]
In den Mitochondrien der kernhaltigen Zellen, z.B. beim Menschen wie ebenso bei wachsenden und sich vermehrenden Hefezellen wird der Luftsauerstoff benötigt, um in der sog. Atmungskette zur effizienten ATP-Produktion die aus dem Wasserstoff H der energiereichen Nahrungsmoleküle wie Stärke bzw. deren Grundbausteinen - Einfachzucker wie Glukose C6H12 O6 - stammenden Elektronen e- und Protonen H+ aufzufangen und abzuführen, um keinen Produktionsstau zu verursachen. Die summarische Reaktion der explosiven Knallgasreaktion 2 H2 + O2 --> 2 H2O + Energie (572 kJ/mol) läuft in den Zellen auf viele kleine Teilschritte aufgeteilt moderiert, also sehr sanft ab. Der Wasserstoff H (= Protonen H+ + Elektron e-) wird an manchen Stellen des aeroben und auch anaeroben Stoffwechsels abgezapft und auf den H- bzw. e- -Überträger NAD+ abgegeben: NAD+ fungiert also als H/Elektronenfänger und Überträger NAD+ + 2 H+/e- --> NADH/H+.
Fazit: Beim aeroben Stoffwechsel wird der Sauerstoff nicht zur Bildung des ausgeatmeten bzw. abgegebenen CO2, sondern in den Mitochondrien
zur Bildung von Wasser verwendet. Die Elektronen zum Betrieb der aeroben Atmungskette und damit zur Synthese von ATP stammen aus der Nahrung. Nahrung wird also nicht "verbrannt", wie das der Ausstoss des
CO2 nahelegt, sondern oxidativ zu CO2 zerlegt und die Energie der
Elektronen in sanften Teilschritten letztlich unter ATP-Gewinn auf Sauerstoff
übertragen.
Beim anaeroben Stoffwechsel wird der z.B. aus der Glukose C6H12O6
abgezapfte H nur zum kleinsten Teil zur ATP-Synthese genutzt ( sog.
Substratkettenphosphorylierung), der Hauptteil wird via Reduktion des
Pyruvats im Alkohol Ethanol C2H5OH gespeichert und somit energetisch
ungenutzt aus der Zelle ausgeschieden.
Abb. 6. Der Wasserstoff H bzw. Protonen H+ und Elektronen e- - Überträger NAD+. NAD+ verhilft zu
Oxidationen, NADH/H+ verhilft zu Reduktionen oder in der zellulären Atmungskette zum ergiebigen ATP-Gewinn.
[Quelle: Markl Biologie, 1. Aufl. (2010), S. 107, modifiziert]


Alkoholische Gärung und unerwünschte Fremdaromen
Bier wird durch die alkoholische Fermentation von Getreide hergestellt. Durch die Keimung der "Stärkespeicher" Gerstenkörner bauen die pflanzlichen Enzyme die enthaltene Stärke zum Doppelzucker Maltose (Malzzucker) ab, die von der Hefe vergoren werden kann. Es entstehen aber nicht nur grossen Mengen an Kohlenstoffdioxid CO2 und trinkbarem Alkohol Ethanol C2H5OH (H3C-CH2-OH), sondern auch diverse Gärungsnebenprodukte (GNP, syn. Bukettstoffe, > 1000 verschiedene Stoffe, cf. Abb. 7), darunter gewünschte und unerwünschte Aromastoffe: Die Biosynthese von Aminosäuren führt zu Nebenprodukten, zu denen sowohl erwünschte Geschmacksstoffe (langkettige Alkohole, syn. höhere Alkohole, Fuselöle) und ca. 90 verschiedene Ester als auch unerwünschte Fremdaromen wie Acetaldehyd C2H4O (H3C-CO-H), Diacetyl H3C-CO-CO-CH3 (C4H6O2) und Schwefelverbindungen gehören.
Die Nebenprodukte können je nach Bierstil zum typischen Geschmack beitragen. Sind diese Nebenprodukte jedoch in zu grossen Mengen vorhanden, entsteht ein unerwünschtes Fremdaroma.
Einige dieser Fremdaromen bilden sich durch die Anwesenheit von Sauerstoff O2, der zum Wachstum und Vermehrung der Hefen notwendig ist, da es sich bei den Hefen um keine echten anaeroben Organismen handelt und einige Zellkomponenten nur durch einen aeroben Stoffwechsel hergestellt werden können. Durch das Vorhandensein von O2 wird aus einem Teil des Pyruvats H3C-CO-COO- (C3H3O3-) --> Acetaldehyd H3C-CO-H (C2H4O) + CO2 gebildet, von dem wiederum eine grosser Anteil in Ethanol H3C-CH2-OH (C2H6O) umgewandelt wird. Aus einem kleineren Anteil des Acetaldehyds entsteht Diacetyl und ein weiterer Teil bleibt erhalten. Sowohl Acetaldehyd als auch Diacetyl verursachen das unerwünschte Fremdaroma (ausführlichere Informationen: cf. Seite Brau-/Bierfehler).
Während der Grossteil des Pyruvats in die alkoholische Gärung eingeht, wird
ein kleiner Anteil für die lebenswichtige Aminosäuresynthese (Bausteine der
Eiweisse!) über Zwischenprodukte des Citratzyklus (Info) abgezweigt. Die α-Keto-
säuren R-CO-COO- des Citratzyklus sind dem Pyruvat analog, wobei die Methyl-
gruppe -CH3 durch eine verlängerte Kohlenstoffkette (R-Gruppe) ersetzt ist.
Eine geringe Menge an α-Ketosäuren wird in langkettige Alkohole umgewandelt,
die wiederum im Bier erwünschte Aromen darstellen (cf. Abb. 6).

Abb. 7. Gärung und Reifung sind begleitet von der Bildung diverser Gärungsneben-produkte, welche Geschmack und Qualität des Bieres deutlich beeinflussen.
Dazu gehören:
-
Jungbukettstoffe (Aldehyde wie Acetaldehyd und Diacetyl, Schwefelverbindungen): werden während Gärung und Reifung wieder abgebaut.
-
Bukettstoffe (Ester, höhere Alkohole): verbleiben im Bier.
[Quelle: Kunze, 2016, S. 402, mod.]
3. SAUERSTOFF: Feind oder Freund des Brauers?
Sauerstoff, Hefen, Hopfen und Bier - eine Geschichte der scheinbaren Widersprüche
1. Was sagt die Hobbybrauliteratur über Sauerstoff?
STOFFWECHSEL: “Die meisten Hefen sind fakultativ anaerob, also nicht auf Sauerstoff
angewiesen. Sie können bei Verfügbarkeit von Sauerstoff, wie die meisten anderen
Lebewesen, ihn für einen oxidativen Energiestoffwechsel nutzen (Aerobe Atmung): Sie
können verschiedene Zucker zu Kohlenstoffdioxid und Wasser oxidieren. In Abwesenheit
von Sauerstoff aber können viele Hefen die Zucker nur zu niedermolekularen Stoffen,
beispielsweise zu Ethanol und Kohlenstoffdioxid (z. B. in der alkoholischen Gärung),
abbauen. Die Zuckeroxidation unter aeroben Bedingungen liefert mehr Energie als die
Vergärung. Deshalb sind die Massenzuwachsrate und die Zellteilungsrate bei oxidativem
Zuckerabbau sehr viel höher als bei der Gärung” [Quelle].
SAUERSTOFF: “Sobald Ihre Würze auf die Gärtemperatur abgekühlt ist und Sie bereit
sind die Hefe anzustellen, sollten Sie die Würze zuerst belüften. Dieser Vorgang liefert
den Sauerstoff, den die Hefe benötigt, um gross und stark zu werden, damit Ihre Würze
vollständig vergoren werden kann.” (Palmer, S. 30, 2020 [Info]).
GÄRUNG: “Belüften der Würze - auch Hefe braucht frische Luft. ..... Hefen vermehren sich im Normalfall durch Sprossung. Die Vermehrung ist deshalb wichtig, weil wir nicht mit der vollen Zellzahl anstellen, sondern sich die Zellzahl im Laufe der Gärung anpasst. Dabei benötigt die Hefe im Normalfall Sauerstoff, weshalb die Würze belüftet wird. .... denn im Grunde braucht die Hefe keinen Sauerstoff.” (Brücklmeier, S. 201, 2018 [Info]).
LAGERUNG: “Es wird sich rund sechs Monate lang halten - und zwar abhängig davon, wie gut es Ihnen gelungen ist, den Sauerstoff während der letzten Gärungsphase und während des Abfüllungsprozesses fernzuhalten.” (Palmer, S. 24, 2020 [Info]).
BIER: “Staatsfeind Nummer 1 - Oxidation. ..... Oxidation ist eine der Hauptursachen für die Bieralterung. Grundsätzlich sollte die Sauerstoffaufnahme des Bieres unbedingt und überall vermieden werden.” (Brücklmeier, S. 244, 2018 [Info]).
SAUERSTOFF: “Sobald Ihre Würze auf die Gärtemperatur abgekühlt ist und Sie bereit sind die Hefe anzustellen, sollten Sie die Würze zuerst belüften. Dieser Vorgang liefert den Sauerstoff, den die Hefe benötigt, um gross und stark zu werden, damit Ihre Würze vollständig vergoren werden kann.” (Palmer, S. 30, 2020 [Info]).
HOPFEN: “Sauerstoff und die dadurch verursachte Oxidation ist neben Wärme der grösste Feind bei der Lagerung von Hopfen.” (Info). “Keep hops away from oxygen. Oxygen is bad, bad, bad for your hops, and limiting exposure to O2 is critical to long-term storage.” (Info).
ALLES KLAR bezüglich Sauerstoff und Bier brauen? Ist Sauerstoff - meistens aus der Luft mit einem Gehalt von 20.95% stammend - nun Freund oder Feind des Brauers bzw. dem Brauvorgang oder dem Endprodukt Bier? Den verschiedenen Aspekten des Sauerstoffs im Zusammenhang mit der Fermentation von Bier soll im Folgenden nachgegangen werden.

Abb. 8. Sauerstoff O2, mit 20.95% ein wichtiger Bestandteil der Luft, ist unentbehrlich für die meisten energieliefernden Reaktionen (z.B. Verbrennungen, Atmung) und einige biosynthetische Reaktionen.
2. Teilaspekt "Sauerstoff und Hefestoffwechsel"
2.1. Hefestoffwechsel 1: Energiebereitstellung
In der Brauliteratur wird meist festgestellt, dass die Brauhefe
Sauerstoff benötigt, mindestens während einer kurzen Zeit
nach dem Anstellen, damit die Hefezellen atmen und wachsen
können. Damit wird angedeutet, dass Hefen Sauerstoff zum
Wachstum und zur Vermehrung in Form der Knospenbildung
benötigen und dazu aerob atmen müssen, bevor die Würze
fermentiert, also anaerob vergärt werden kann.
Grundlagen des Stoffwechsels für alle Organismen: Lebens
basiert auf einem Kohlenstoff-Stoffwechsel und einem Energie-
Stoffwechsel. Während Pflanzen (und wenige Bakterienformen)
ihre Lebensenergie aus der in C-Verbindungen wie Glukose
C6H12O6 oder Stärke (C6H12O6)n fixierten Sonnenenergie
holen können (Prozess: Fotosynthese, sog. autotrophe
Lebensweise), sind fast alle anderen Lebewesen wie Tiere,
Mensch und auch die Hefen auf energiereiche C-haltige Stoffe
(“Lebensmittel”) angewiesen, um ihren Bau- und Energie-
stoffwechsel betreiben zu können (Prozesse: aerobe Atmung,
anaerobe Fermentation [Gärungen], sog. heterotrophe
Lebensweise).
Der allen heterotrophen Lebewesen gemeinsame Stoffwechsel oder Metabolismus beruht zunächst auf dem Katabolismus, dem Abbau der energiereichen C-haltigen Stoffe wie Stärke, verschiedener Zuckerarten, Fette und Eiweisse mit den daraus resultierenden Aufbau-Molekülen bzw. Molekülgruppen und dem biologischen Hauptenergieträger ATP (Adenosin-tri-phosphat). Aus den C-haltigen Bausteinen und dem Energieträger ATP werden im Anabolismus, dem Aufbaustoffwechsel die Grundbestandteile der Zellen wie z.B. Membranlipide (Abgrenzung) und Membranproteine (Transportfunktionen, u.a.) gebildet (Biosynthesen) [gute Verständnis erweiternde Abb. hier].
Hefen nun sind mit der einmaligen Fähigkeit ausgestattet, dass sie zwei Hauptstoffwechselwege der ATP-Energiegewinnung besitzen, nämlich sowohl die sauerstoff-abhängige aerobe Atmung (Zellatmung) als auch die sauerstoff-unabhängige anaerobe Gärung.

Abb. 9. Der Stoffwechsel kann grob in zwei Hauptreaktionswege gegliedert werden, in den Abbaustoffwechsel Katabolismus und den Aufbaustoffwechsel Anabolismus.
Atmung ist nicht nur die Aufnahme von Sauerstoff, sondern vielmehr ein komplexer biochemischer Prozess, der primär in den Atmungszentren der Zellen, den Mitochondrien abläuft und z.B. Traubenzucker (Glukose) vollständig oxidiert zu Wasser H2O, Kohlenstoffdioxid CO2 und dem energiereichen Hauptprodukt ATP, das eine Art energetisches Kleingeld für alle energieabhängigen biochemischen Reaktionen in der Zelle/Gesamtorganismus unentbehrlich ist.
Zellatmung im Überblick: verläuft in drei grossen Reaktionskomplexen ab
- 1. Glykolyse (“Zuckerauflösung”) im Cytoplasma der Zelle –> liefert nur gerade
2 Energieeinheiten ATP + 2 Pyruvat C3 (Brenztraubensäure)
- 2. Citratzyklus im Innenraum der zellulären Atmungszentren (Mitochondrien)
–> 2 ATP + zahlreiche Bausteine für Biosynthesen
- 3. Atmungskette an innerer Mitochondrienmembran –> 28 ATP
Summengleichung: Glukose C6H12O6 + 6 O2 + 6 H2O —> 6 CO2 + 12 H2O + 32 ATP
(=vollständige Oxidierung von Glukose)
Gärung (engl. fermentation) ist eine Energiefreisetzung ohne Sauerstoff, aber mit wenig
Gewinn von ATP und energiereichen Endprodukten wie Alkohol (alkoholische Gärung)
oder Milchsäure (Milchsäuregärung). Gärungen beginnen wie die Zellatmung mit der
Glykolyse, wobei nur 2 energiereiche ATP-Moleküle gewonnen werden. Das C3-Pyruvat
wird in 2 Reaktionsschritten zu Ethanol (“Alkohol”) umgewandelt.
- 1. Glykolyse (“Zuckerauflösung”) im Cytoplasma der Zelle –> liefert nur gerade 2
Energieeinheiten ATP + 2 Pyruvat C3 (Brenztraubensäure)
- 2. Ethanolbildung –> 2 C2H6O (CH3-CH2-OH) + 2 CO2
Summengleichung: Glukose C6H12O6 –> 2 C2H6O (CH3-CH2-OH, C2H5OH) + 2 CO2 + 2 ATP
Teiloxidierung von Glukose, energiereiches Endprodukt Ethanol (Alkohol)
Die Hefe hat also einen Luxusstoffwechsel, sie kann sowohl mit Sauerstoff (aerob) als auch ohne
Sauerstoff (anaerob) leben. Sie zeigt aber eine ausgesprochene Tendenz zur Gärung und atmet
meist nur dann, wenn sie 1. wichtige Zellbausteine zum Wachstum und Zellvermehrung
synthetisieren muss, und 2. die Zuckerkonzentration sehr gering und gleichzeitig Sauerstoff
vorhanden ist.
Hefe kann mehrere Zuckerarten vergären (Diauxie), sie ist allgemein sehr flexibel bezüglich
verwertbarer C-Substrate; so kann sie z.B. sogar Ethanol verwerten (allerdings nur bei
Abwesenheit von vergärbaren Zuckern –> Essigsäure).
Hefe hat also letztlich 3 Stoffwechselwege zur Verfügung:
1. Anaerobe Gärung bei Vorhandensein von Zucker und Abwesenheit von Sauerstoff
2. Aerobe Atmung: bei geringer vergärbarer Zuckerkonzentration und Anwesenheit von
Sauerstoff
3. Aerobe Gärung bei Vorhandensein von viel Zucker und Sauerstoff.
Abb. 10. Stoffwechselzusammenhänge: Glykolyse (--> Pyruvat C3), Gärungsabzweigung (Alkoholische Gärung, Milchsäuregärung), Citronensäurecyclus, Atmungs-kette.
Involvierte Moleküle: Glucose C6, Pyruvat C3, Acetyl-CoA C2, O2, CO2, H2O. Energieträger ATP und Wasser-stoffüberträger NADH und FADH2 (cf. Abb. 5+6).

Was steuert wie den Hefestoffwechsel?
Der sog. Pasteur-Effekt geht auf ein 1861 von Louis Pasteur entdecktes Phänomen zurück. Er hat beobachtet, dass Hefen unter anaeroben Bedingungen mehr und schneller D-Glucose verbrauchen als unter aeroben Bedingungen. Die Hefen verarbeiten also unter anaeroben Bedingungen weiter Zuckermoleküle, weit mehr als unter aeroben Bedingungen, um die Energieverluste (- 32 ATP) zu kompensieren. Gleichzeitig wird somit auch viel mehr Ethanol produziert.
Der Crabtree-Effekt, bezeichnet nach dem englischen Biochemiker Herbert Grace Crabtree beschreibt das Phänomen, dass die Hefeart Saccharomyces cerevisiae auch unter aeroben Bedingungen Ethanol produzieren kann und nicht Biomasse/Zellsubstanz zum Zellaufbau oder zur Zellvermehrung via den unter aeroben Bedingungen sonst typischen Stoffwechselweg des Citratzyklus synthetisiert wie das sonst andere Hefearten aerob tun, z.B. Kluyveromyces spp. Voraussetzung dazu ist eine “hohe” äussere Glukosekonzentration. Neben Glukose, das bereits ab 100 mg Glukose pro Liter Nährmedium den Crabtree-Effekt bewirkt, lösen auch die anderen vergärbaren Zuckerarten wie Fruktose, Maltose, Saccharose und teilweise Galaktose diesen Effekt aus. Eine Würze mit 1.040 OG (bzw. 10 oP) ist 10%ig zuckerhaltig! Diese sehr hohen Zuckerkonzentrationen unterdrücken also fast vollständig die aerobe Atmung in der Anstellwürze und forcieren die Gärung!
2.2. Hefestoffwechsel 2: Biosynthese von Zellmembrankomponenten
Die eigentliche Aufgabe des Sauerstoffs in der belüfteten Anstellwürze - nicht Atmung/Energiebereitstellung, sondern Förderung spezifischer Biosynthesen!
Die Hefe braucht letztlich keinen Sauerstoff, sie überlebt auch ohne. Sie kann auch unter strikt anaeroben Bedingungen wachsen. Trotzdem wird in der Brauliteratur und aufgrund der Brauererfahrungen immer wieder auf die Notwendigkeit von Sauerstoff für eine gesunde Fermentation hingewiesen. Welche genaue Rolle spielt also Sauerstoff für einen kräftigen und gesunden Gärverlauf?
Viele Untersuchungen zeigen eindeutig auf, dass Hefe Sauerstoff verbraucht wann immer er verfügbar ist, auch während des Gärvorgangs. Hefezellen absorbieren sehr rasch praktisch den gesamten zur Verfügung stehenden Sauerstoff. Hefen verbrauchen den überwiegenden Anteil des verfügbaren Sauerstoffs für biosynthetische Reaktionen, nicht für die Atmung bzw. Energiegewinnung. Der gelöste O2 in der Würze wird durch frisch angestellte Hefen sehr rasch von der Sättigungskonzentration von ca. 14 mg O2/L auf Null reduziert, meistens innerhalb von 30 Minuten unter idealen Bedingungen. Warum? Hefen benötigen diesen Sauerstoff für die Biosynthese von Membranbestandteilen, welche wiederum die Voraussetzung für rasches Wachstum und Zellvermehrung zur Erreichung einer hohen Zelldichte notwendig ist. “Oxygen has profound influence on the activity of yeasts and particularly on yeast growth. Certain yeast enzymes react only with oxygen, and oxygen cannot be replaced by other hydrogen acceptors.This applies to the oxygenases involved in the synthesis of unsaturated fatty acids and sterols which are vital elements of the cell membranes.Without a supply of these lipids,the cells cannot reproduce and their viability is lessened” (Quelle). Es sind dies vor allem sog. Sterole (besonders Ergosterol) und ungesättigte Fettsäuren. Ohne diese Membrankomponenten in der Würze ist das Zellwachstum limitiert (Info), oder anders formuliert, ohne Sauerstoff zu Beginn der Gärung limitieren diese fehlenden Membrankomponenten Wachstum, Vitalität und Zellvermehrung.
Synthese von Sterolen (Sterine) und ungesättigten Fettsäuren
Während Zellwände von Zellen aus Kohlenhydraten
bestehen (Pflanzen: zum grössten Teil aus Zellulose,
letztlich einem Vielfachzucker aus Glukose [Info], Hefen:
Glucane und Mannane [Info1, Info2], ebenfalls aus
Glukose-Bausteinen), ist die Synthese von Sterolen
aus Isopren-Einheiten (Info1, Info2) und die der
ungesättigten Fettsäuren aus Acetyl-Coenzym A
(aktivierte Essigsäure, Info) recht komplex. Sauerstoff
spielt im Fettstoffwechsel eine wichtige Rolle. Zunächst
nimmt die Hefe die in der Würze vorhandenen Fettsäuren
auf. Sie ist aber auch in der Lage, unter Sauerstoffaufnahme
selbst essenzielle Fettsäuren zu bilden. Diese Bildung
beginnt beim Pyruvat (Brenztraubensäure) und führt
über Acetyl-Coenzym A. Die Hefe verfügt aber auch die
Stoffwechselfähigkeit, ungesättigte Fettsäuren zu bilden.
Ebenfalls unter Sauerstoffaufnahme wird ein Teil der
Fettsäuren in ungesättigte Fettsäuren umgewandelt.
Sterine (von gr. fest, starr, hart, wegen der relativ
starren Molekülstruktur), syn. Sterole, sind eine Gruppe
von Membranlipiden und biochemisch wichtige Bestandteile
der Zellmembran. Sterine sind wichtig für die Membran-
eigenschaften wie Fluidität, Elastizität und Permeabilität.
Hefen können die Sterine entweder aus der Anstellwürze
aufnehmen oder selbst herstellen. Sauerstoff wird nun auch
für die Biosynthese der Sterine benötigt.
Dieser Synthesestoffwechsel zu Beginn der eigentlich
anaeroben Gärung kann als “respiratorische Fermentation”
bezeichnet werden, d.h. aerobe Atmungsprozesse und Gärung
laufen simultan ab, obgleich die Gärungsprozesse dominieren.
Der respiratorische Stoffwechsel (Atmungsstoffwechsel) zapft
höchstens etwa 1-2% der gesamten Glukose für die
biosynthetischen Stoffwechselpfade der Sterol- und Fettsäuren-
synthesen ab. Der damit verbundene Sauerstoffbedarf an
gelöstem Sauerstoff in der Würze ist also kein Atmungssauerstoff
zur Energieproduktion (d.h. keine aerobe Atmung für die ATP-Synthese)
noch wird er direkt für die Hefeknospung benötigt. Die Membran-
komponenten sind aber für vitale Membranfunktionen, primär
Kontrolle der Stoffflüsse über die Membran in und aus der Hefezelle
verantwortlich (Phospholipide mit eingebetteten Transport-Proteinen),
damit für die Vitalität der Hefezellen allgemein. Geschwächte Hefezellen
sind für eine Vielzahl von Problemen verantwortlich, so z.B. Intoleranz
gegenüber Alkohol (Alkohol schwächt dann die Zellen massiv), verlangsamte oder stockende Gärung und zahlreiche Bierfehler/Fehlaromen.

Abb. 11. Bau einer Zellmembran der Hefe, stark vereinfachte Darstellung.
Ungesättigte Fettsäuren (UFS) in der Lipiddoppelschicht und Sterine sind wichtige Bausteine der Hefezellmembran. Sterine wie z.B. Ergosterol (Abb. 11) kommen in geringerer Konzentration als die Fettsäuren vor, aber sie sind sehr wichtig für die Fluidität (Fliesseigenschaft) der Membranen und damit verantwortlich für deren Elastizität und Permeabilität (Stoffdurchlässigkeit). Proteine können eingelagert oder integral (membran-durchstossend) sein und übernehmen wichtige Funktionen, z.B. selektiver Transport von Zuckerarten, Aminosäuren (--> Proteinaufbau), Mineralstoffen, Signalempfang u.a.

Abb. 12. Struktur des Sterols Ergosterol.
Vitalität, Viabilität, Gärkraft, Vermehrung durch Zellknospung, Bau der Zellorganellen
(z.B. Mitochondrien [Zellkraftwerke, Zellatmung], Kernmembran, Vakuolenmembran,
u.a. membranumhüllte Organellen: siehe Abb. 13, auch Bau der Hefezelle) und viele
weitere stoffwechselgeprägte Phänomene von Hefezellen sind von den Eigenschaften
der Biomembranen (syn. Zellmembranen) beeinflusst. Biomembranen sind also das
wichtigste Verpackungsmaterial der Zellen, das den Zellinhalt (wässrige Lösungen)
möglichst gut zusammen hält, aber selbst nicht wasserlöslich ist. Zudem sollten sie
eine flexible Bauweise erlauben, damit Zellen und deren Zellorganellen bei Bedarf
vergrössert, verkleinert oder umgebaut werden können.
Bei der Vermehrung/Sprossung und Wachstum von Hefen muss eine enorme
Syntheseleistung an Zellsubstanzen erbracht werden, insbesondere auch an
Membranen: das sind unter Berücksichtigung der membranumhüllten
Zellorganellen riesige Oberflächen. Kunze berechnet aus der Oberfläche einer
einzelnen Hefezelle von ca. 150 μm2 eine Totalkontaktfläche von 9 bis 10 m2
bei 10 g abgepresster Hefen (Info, S. 10).
Welcher Brauer wünscht sich nicht vor Vitalität stotzende Hefen mit hoher Gärkraft?
Es lohnt sich deshalb, auch einmal einen vertieften Blick zum Verständnis gewisser
Eigenheiten der Hefezellen in die Biochemie der Biomembranen zu werfen. Auch
daraus können wichtige Erkenntnisse für die Braupraxis gewonnen werden.
Hinweis: Der allgemeine Bau der Biomembranen wird mit den Abb. 14 bis 16
erläutert: Fettsäuren --> Phospholipide --> P-Lipiddoppelschichten.
LIPIDE, BIOMEMBRANEN Vertiefende Informationen

Abb. 13. Bau einer Hefezelle. ROT: Membranstrukturen
1: Sprossnarbe. 2: Plasmaeinbuchtung. 3: Plasmamembran = Zellmembran. 4: Zellwand. 5: Mitochondrium (Zellatmung). 6: Lipid-granula (Lipidspeicher). 7: Mitochondrium. 8: Golgikomplexe ("Endmontage" von Eiweissen"). 9: Vakuole (Speicher, Stoffaufspaltung, Abfälle). 10: Polyphosphatkörner (Energiespeicher). 11: Endo-plasmatisches Reticulum (verzweigtes Membrankanalsystem mit verschiedensten Stoffwechselfunktionen, Info). 12: Kernmembran.
13: Zellkern (Erbinformation). 14: Kernkörperchen (Info).
[Quelle: Annemüller, Info, 2020, S. 142; mod.]
Bau der Hefezellwand und Zellmembran

Abb. 14. Bau der Hefezellwand und Hefezellmembran.
Die Zellwand ist dreischichtig. Die äussere Schicht ist plastisch, es folgt eine Stützmembran aus Glucane und Mannane sowie Chitin. Die feine Innenmembran, die Zellmembran besteht aus Proteinen (Eiweisse) und Phospholipiden.
Mannoproteine: Verbindungen zwischen Eiweissen und Polysacchariden (Mehrfachzucker) als
natürlicher Bestandteil der Hefezellwand
Glucane: Polysaccharide, die in vielen pilzlichen Zellwänden als struktureller Baustein mit
festigender Wirkung enthalten ist
Mannane: verzweigte Polysaccharide aus D-Mannose in Hefezellwand
Chitin: Polysaccharid, das der Strukturbildung dient und u.a. in der Zellwand von Pilzen
Lipide: Gesamtheit der Fette und fettähnlichen Substanzen
Phospholipide: komplexe Lipide, welche eine Esterbindung mit Phosphorsäure aufweisen. Sie sind
Hauptbestandteil aller Zellmembranen und gehören daher zu den Membranlipiden
integrale
Proteine: Membranproteine, die zum Teil in eine Membran eingebettet sind oder diese ganz
durchdringen, z.B. mit Transportfunktionen (Bsp. Maltosetransporter, Glukose-
transporter)
Wichtige Grundbausteine (=Makromoleküle) der Zellmembranen
Lipide: sind Kohlenwasserstoffverbindungen und bestehen aus unterschiedlichen Gruppen wie
- Fette (aus Glycerin + 3 Fettsäuren): cf. Abb. 15
- Phospholipide (cf. Abb. 16)
- Sterole (starres Gerüst aus 4 Kohlenstoffringen, Bsp.
Cholesterol, syn. Cholesterin) und Steroide (C-Ringsystem
mit 3 Sechser- und 1 Fünferring: cf. Abb. 12).
Abb. 15. Bausteine der Fette: Fettsäuren sind Moleküle mit einer langen unverzweigten Kohlenwasserstoffkette -CH2- und einer Carboxylgruppe (-COOH).
Bsp. gesättigte* Fettsäure Stearinsäure C18: H3C(CH2)16COOH
ergeben dicht gepackte Fettsäureschwänze in Membranen (*: gesättigt, weil an den C-Atomen der Kohlenwasserstoffkette alle 4 Bindungsstellen mit 2 C und 2 H-Atomen besetzt sind [-C-C-C- und -CH2].
Bsp. ungesättigte Fettsäure (UFS) Linolsäure C18:
H3C(CH2)4CH=CHCH2CH=CH(CH2)7COOH =: Doppelbindung -HC=CH-, ungesättigt, weil dort noch H-Atome am C "fehlen"
UFS haben einen niedrigeren Schmelzpunkt und durch Doppel-bindungen ergeben eine geknickte Schwanzstruktur, dadurch eine weniger Dichte Packung in Membranen und eine bessere Elastizität/ Fluidität (--> cf. Abb. 17).
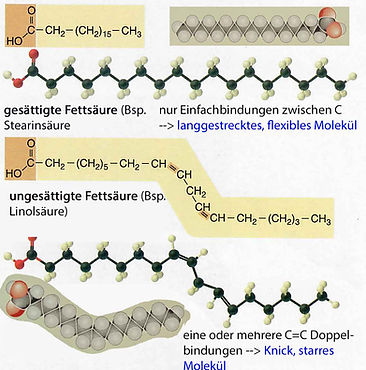
Abb. 16. Bau der Zellmembranen aus Phospholipiddoppelschichten.
Die Grundstruktur besteht aus einer Lipiddoppelschicht aus Phospholipidmolekülen. Darin eingesenkt oder ganz durchlaufend sind verschiedenste Proteine, meist für Transport- funktionen in und aus der Zelle (cf. Abb. 11). Info Lipiddoppelschicht. Quelle: Sadava, Life (2010), mod.

Abb. 17. Elastizität/Fluidität von Biomembranen hängt von den in den Lipiddoppelschichten eingebauten Fettsäuren ab.
Membranen mit Phospholipiddoppelschichten mit ungesättigten Fettsäuren sind weniger dicht gepackt, weisen dadurch eine flexiblere "flüssigere" Struktur auf und lassen Moleküle (z.B. Nährstoffe) eher durch als solche mit nur gesättigten Fettsäuren.

Fettsäuren sind also für den Bau der Biomembranen und damit für zahlreiche Zellstrukturen und Funktionen essenziell. Wo aber braucht es jetzt konkret Sauerstoff für die Biosynthese der Lipide, spezifisch z.B. für ungesättigte
Fettsäuren oder Sterole?
Die Bildung von Doppelbindungen im Lipidschwanzteil gesättigter Fettsäuren
und in Squalen (C30H50, ein Alken mit 6 Doppelbindungen), dem Baustein für
Sterole ist ein Mehrschrittprozess, der Sauerstoff O2 benötigt. Abb. 18 zeigt die
stark vereinfachte Reaktion der Bildung einer Doppelbindung. Die Sterinsynthese
ist komplex. Kurz zusammengefasst die wichtigsten Reaktionsschritte:
1 Glykogen (“tierische Stärke”) –> 2 Acetyl-CoA (aktivierte Essigsäure) –> 3 Squalen
+O2 –>4 2,3-Epoxysqualen –> 5 Lanosterol –> andere Sterole wie z.B. 6 Ergosterol (Abb. 18).
Die ebenfalls O2-abhängige Biosynthese einiger ungesättigter Fettsäuren ist schematisch in Abb. 20 dargestellt.

Abb. 18. Bildung einer Doppelbindung in ungesättigten Fettsäuren oder im Sterol-baustein Squalen unter O2-Verbrauch.

Abb. 19. Sterinsynthese: führt über zahlreiche Reaktionsschritte und Zwischenprodukte.
1: Glykogen (Glukose-Speicher). 2: Acetyl-CoA ("aktivierte Essigsäure"). 3: Squalen (C30H50). 4: 2,3-Epoxysqualen (C30H50O). 5: Lanosterol (C30H50O). 6: Ergosterin (C28H44O).
Die Umwandlung von 2 nach 3 umfasst 10 enzymatische Reaktionsschritte, von 3 nach 6 sind weitere 10-12 enzymatische Reaktionsschritte notwendig. Die Sterine werden dann in die Zellmembran und Organellen-membranen eingebaut. Der Sauerstoffbedarf ist mit O2 markiert.
Detaillierter Reaktionsverlauf 3 --> 6: cf. hier.

Abb. 20. Biosynthese ungesättigter Fettsäuren.
Beispiel Arachidonsäure aus Linolsäure [Abb. mod., Quelle]

Abb.21. Sauerstoff wirkt sich an spezifischen Stellen im Brauprozess positiv wie auch (meist) negativ aus.
2.3. Zusammenfassung/Konsequenzen "Hefestoffwechsel und Sauerstoff"
-
Energiegewinnung: Hefe atmet selten, hohe Zuckerkonzentrationen unterdrücken
die aerobe Atmung und fördern die anaerobe Gärung.
-
Sauerstoffbedarf: Der Sauerstoffbedarf der Anstellhefe ist durch die Fettsäure-
bildung und Sterinbildung verursacht und nicht durch aerobe Atmung. Beide
Lipidstoffklassen sind essentiell für die Membranausbildung, damit für Wachstum,
Vermehrung durch Knospenbildung der Hefezellen und somit auch für ihre
Vitalität (Hefeaktivität, Gärleistung/Gärkraft) und hohe Viabilität (Lebensfähigkeit,
wenig Totzellen in der Anstellhefe).
-
Anstellwürze: Eine optimale Anstellwürze enthält Membrankomponenten wie
ungesättigte Fettsäuren und Sterine (meistens aus toten Hefezellen oder
Hefezellwänden/Membranen gewonnen). Wenn belüftet soll der Sauerstoffgehalt
nie 6-8 ppm O2 überschreiten. In kühler Würze wird mehr Sauerstoff gelöst als in
warmer Bierwürze.
-
Belüftung: Die Herstellung und sterile Belüftung einer Starterkultur ist optimal und
einer starken Belüftung der Anstellwürze vorzuziehen. Von der frischen Starterkultur
sollte nur der Hefeanteil (Hefesediment, nicht der flüssige Überstand) in die Anstellwürze überimpft werden, um eine evtl. negative Geschmacks-
beeinflussung zu vermeiden.
-
Anstellzellzahl: Eine optimale Anstell-Hefezahl (weder zuwenig noch zuviele Hefzellen) fördert die Bierqualität und minimiert Bierfehler.
2.4. PRAKTISCHE KONSEQUENZEN für die Primärgärung
Mangelnder Sauerstoff zu Beginn der Gärung kann negative Auswirkungen haben wie
- mangelnde Angärung
- längere Gärdauer (bis zu 3 oder mehr Tagen)
- gestresste Hefen, verminderte Viabilität der abgeernteten Hefen
- stockender Extraktabbau, unvollständige Gärung
- mangelnde Nachgärung, ungenügender Vergärungsgrad
- Probleme mit der Bierqualität, z.B. Bierfehler wie exzessive Esterbildung
1. Lipidzugaben statt Sauerstoff bzw. als Ergänzung
Theoretisch könnte der Sauerstoffbedarf frisch angestellter Hefen durch Zugabe benötigter
Fettsäuren und Sterine reduziert oder gar eliminiert werden, z.B. in Form der Zugabe geeigneter
organischer Hefenährstoffe ([Info]. Geeignete Produkte: z.B. Lallemand Go-FERM-Protect Evolution,
cf. Baldinger, Info. SIHA PROFERM Bio yeast nutrient, Info1, Info2), SIHA Hefenährstoff-Navigator,
cf. Eaton Deutschland; allgemein tote Hefezellen (z.B. Yeast Hulls) oder auch Kalttrub (evtl. Nachteil
von Geschmacksfehler, Info).
2. Belüftete Starterkultur statt Würzebelüftung
Eine ausreichende Zahl vitaler Anstellhefen aus einer gerührten und steril mit Luft belüfteten
Starterkultur kann eine Belüftung der Anstellwürze ersetzen oder aber ergänzend sich positiv
auf den Start und den Verlauf der Primärgärung auswirken.
Eine Würzebelüftung sollte 8 - 10 ppm O2 nicht übersteigen, auch weniger O2 kann je nach Hefestamm optimal sein (cf. auch Info hier). Ideal wäre eine genaue O2-Bestimmung mittels einer O2-Elektrode, damit keine Unter- noch Überbelüftung stattfindet (cf. Info1, Info2).
Rehydratisierte Trockenhefen benötigen i.d.R. weniger Sauerstoff, da sie während ihrere Kultivierung erhebliche Glykogen- und Lipidreserven anlegen konnten. Daher genügt das Schütteln des Starters und Eingiessen in die Anstellwürze.
Auswirkungen des gelösten Sauerstoffs:
zu wenig: Esterbildung +, Bildung Fuselalkohole -
zu viel: Esterbildung -, Bildung Fuselalkohole +, Problem mit Acetaldehyd
Hinweis:
Faustregel für optimale Anstellzellzahl ca. 1 Million aktiver Hefezellen pro mL
Würze und pro Grad Plato, noch vereinfachter ca. 5 bis 10 Mio. Zellen/mL Würze.
Differenzierte Berechnung der Anstellzahlen: cf. “Bestimmung der optimalen
Anstellzellzahl (Hefegabe)” > Braulabor 19).
Würzebelüftung: siehe Braulabor 21 “Beimpfung der Bier-Anstellwürze” > Anleitung
hier (Schritt 5 “Belüftung und Hefeeinmischverfahren” S.5-6). Bei einer Überbelüftung,
insbesondere mit reinem Sauerstoff können unerwünschte Substanzen entstehen
wie VDKs (vicinale Diketone, Info), Diacetyl, Oxosäuren, Fuselalkohole und abgeleitete
Esterverbindungen.
3. Belüftung kann bierstiltypisch sein
Bei gewissen Bierstilen ist es sogar empfehlenswert, die kalte Anstellwürze nicht zu
belüften, z.B. bei hohen Platowerten, die zur stärkeren Esterbildung tendieren (Bsp.
Strong Scotch, OG 1.08-1.10/19-24 °P). Erwünschte oder unerwünschte Esterbildung
kann also direkt durch die Belüftungsdauer der Würze beeinflusst werden.
Andrerseits muss bei Bieren mit sehr hoher Stammwürze > 22 °P (OG 1.083) nach
etwa 12 bis 18 Stunden nach dem Anstellen eine zweite Dosis mit reinem Sauerstoff
gegeben werden (> 12 ppm O2 bis 17 ppm).

Abb. 22. Belüftung der Anstellwürze.
li: mit reinem O2 aus O2-Druckflasche
re: mittels Aquariumpumpe, Sterilfilter und porösem Belüftungsstein. [Info1 Info2]

Abb. 23. Go-Ferm-Protect Evolution ist eine Hefenahrungs-zusatz zur Hydration oder Starterkulturen.
Es enthält u.a. Vitamine und Sterole und kann somit die Initial-belüftung beim Anstellen zu Beginn der Primärgärung substituieren.
3. Teilaspekt "Sauerstoff und Braurohstoffe, Bier und Brauprozesse"
3.1. Kurze Übersicht
"Staatsfeind Nummer 1 - Oxidation" - so ist ein Kapitel in der "deutschen Bierbibel" von Brücklmeier überschrieben (Brückelmeier, 2018, S. 244). Sauerstoff ist zwar nicht sehr reaktionsfähig und chemische Reaktionen mit Sauerstoff werden im engeren Sinne auch als Oxidationen bezeichnet (Info). Oxidationen können sowiohl die Brauausgangssstoffe wie Malze und Hopfen beeinflussen wie auch das Produkt Bier direkt. Oxidation ist nämlich eine der Hauptursachen der Bieralterung. Deshalb sollte die Sauerstoffaufnahme des Bieres während des gesamten Brauprozesses, also vom Schroten bis zum Biergenuss in der Kehle - mit eben der Ausnahme der Anstellwürze bzw. Starterkultur - minimiert werden!
Sauerstoff O2 kommt in der Luft zu rund 21 Volumen-Prozent vor, d.h. was immer wir auch
machen, Sauerstoff umgibt uns ständig, ist unser Dauerbegleiter. Ob im "heissen Teil" des
Brauens ( -->Herstellung der Anstellwürze), dem "kalten Teil" der Gärungs- und Reifungsphasen,
bei den Transfers von einem Gärbehälter in einen anderen Behälter, bei der Abfüllung der
Flaschen, der Reifung und Lagerung - Sauerstoff ist einfach da!
Elementarer, gasförmiger Sauerstoff ist zwar relativ reaktionsträge, viele Reaktionen finden bei
Normalbedingungen gar nicht oder nur langsam statt. Zur Reaktion werden also entweder
eine hohe Aktivierungsenergie oder sehr reaktive Radikale benötigt. Diese Barriere kann aber
durch Temperaturerhöhung, Licht oder Katalysatoren überschritten werden. Beim Brauprozess
sollte also beachtet werden:
-
möglichst kein Zutritt von Luft (Sauerstoff) zu den verschiedensten Phasen des Brauprozesses
(Bsp. Bierwürzen in keiner Phase "plätschern" lassen, Flaschen möglichst voll auffüllen)
2. tiefe Temperaturen erschweren Oxidationen (Bsp. kalt lagern 0 °C - < 10 °C)
3. Zeit ist ein wichtiger Parameter, gerade bei langsamen Reaktionen, ebenso der Ausschluss
von Licht (beide Faktoren wirken während Reifung und Lagerung).

Abb. 24. Sauerstoff O2 umgibt uns überall und immer. Unerwünschter Sauerstoffeintrag während Umfüllprozessen und Lagerung von Bier.
3.2. Sauerstoff und Braurohstoffe, Bier und Brauprozesse
MALZ und Sauerstoff.
Durch das Schroten des Malzes werden die Körner aufgebrochen
und dadurch vergrössert sich die Oberfläche deutlich. Der Kontakt
mit der Luft und somit dem Luftsauerstoff wird intensiver durch
diese vergrösserte Oberfläche und den Wegfall der schützenden
Hülle der Fruchtschale (cf. Abb. Gerstenkorn hier). Geschrotetes
Malz verliert nach dem Mahlen durch Oxidationsprozesse
zunehmend Stärke, Eiweiss und vor allem die für die Alkoholbildung
wichtigen Enzyme.
Abhilfe: Unmittelbar vor dem Maischen schroten.
Das geschrotete Malz sollte nie länger als etwa eine Woche
gelagert werden, ausser vakuumiert und tiefgefroren.
HOPFEN und Sauerstoff.
Hopfen wird in 3 Produktformen angeboten: getrocknete ganze
Hopfendolden (Rohhopfen), Hopfenpellets und Hopfenextrakt.
Von den Hopfeninhaltsstoffen Bitterharze, Hopfenöle, Polyphenole,
Proteine und Lipide sind vor allem die letzteren mit den ca. 3%
freien Fettsäuren für die Hopfenalterung verantwortlich. Bei
unsachgemässer Lagerung, also weder licht- sauerstoffgeschützt
noch kalt bis tiefgefroren verliert der Hopfen durch Oxidationen
rasch die Aromen und Bitterharze, bildet vermehrt freie Fettsäuren
(3% –> 20%) und entwickelt unangenehme Aromen nach Knoblauch
und Käse-/Schweissfuss.
Abhilfe: Rohhopfen sofort verbrauchen (< 1-2 Tage), oder rasche
Trocknung und Vakuumverpackung oder Verpackung mit Inertgas
wie Kohlenstoffdioxid CO2 oder Stickstoff N2, Lagerung gekühlt
oder besser noch tiefgefroren und lichtgeschützt (cf. Abb. hier).
BIERalterung durch Oxidation.
Darunter wird vereinfacht die Veränderung der Geschmacksstabilität vom idealen Zustand des frisch vergorenen
und gereiften Bieres verstanden.
Zwei Defizite zeichnen alterndes Bier aus, nämlich 1. der Verlust sog. positiver Biereigenschaften, und
2. der Alterungsgeschmack.
Von den oxidativen Eigenschaftsverluste betroffen sind 1. Hopfenöle (–> Hopfenblume), 2. Melanoidine
(–> Aromen, insbes. dunkle Biere), 3. Kolloide (–> z.T. Vollmundigkeit) und 4. Iso-Alphasäure (–> feine
Hopfenbittere).
Der Alterungsgeschmack wird zurückgeführt auf verschiedene chemische Stoffe wie Schwarze Johannisbeere/
Katzenurin (–> zu grosser Flaschenkopfraum), Oxidation von Hopfenölen/Polyphenole/Proteine (–> nach-
hängende Bitterkeit), Strecker-Aldehyde (–> brotig, karamelig/süsslich, honigartig, mandelartig), Oxidation
von Fettsäuren in alten Flüssigmalzextrakten (–> Trans-2-Nonenal: fahl, abgestanden, seifig, Pappkarton/
Altpapier), Diacetyl (aus Hefe-Acethydroxysäuren + O2 [Butter/Popcorn]), Lösungsmittel aus Kunststoffen
(z.B. PVC –> lösungsmittelartiger Beigeschmack).
Aber auch nicht-oxidative Veränderungen können zum Altersgeschmack beitragen (z.B. feuchte Pappe,
Maillard-Reaktionen, Veresterung).
Mehr Info: siehe auch unter Brau-/Bierfehler > 5. Fehlaromen. Info1 Info2.
Auch die Bierfarbe kann von Oxidationen betroffen sein (Polyphenole + O2, Kohlenhydrate + Proteine +
Hitze –> Maillard-Reaktionen –> dunkle Farbe, u.a.).
Abhilfe: Allgemein gilt möglichst geringer Kontakt mit Luft (–> enthält ja 20.95% Sauerstoff), möglichst
tiefe Temperaturen (–> Verlangsamung chemischer Reaktionen, z.B. Oxidationsreaktionen) und lichtgeschützt.
Konkrete O2-minimierende Massnahmen:
-
Antioxidativ wirksame Substanzen vermehren: + Melanoidine (dunkle Malze, Caramalze).
-
Hoher Polyphenolgehalt (richtige Hopfenwahl).
-
Hoher Schwefeldioxid (SO2)-Gehalt: f(Hefestamm!), Sulfat-Konzentrationen: SO4 2- durch Calciumsulfat CaSO4: bis 150 mg/L, Ales bis 300 mg/L).
-
Eisen- und Kupferionen meiden.
-
Hohe pH-Werte vermeiden (günstig: pH 5.2 - 5.6 in Maische, pH 5.4 in Würze, pH 5.2 in Würze 10 min vor Kochende).
-
Wärmeeinwirkung reduzieren: optimaler Wärmeübergang in Sud, Würzekochen max. 90 min.
-
Vermeidung tiefer Einmaischtemperaturen (–> Reduktion Aktivität hitzestabiler Malz-Oxidasen –> weniger Polyphenoloxidationen).
-
Ganzen Brauprozess optimieren: z.B. Maischen, Läutern, Ausschlagen der Würze möglichst O2-arm (z.B. durch Begasung mit CO2/N2), Prozesszeiten minimieren (–> kürzere Kontaktzeiten mit Luftsauerstoff), Flüssigkeiten nicht plätschern, Einleiten/Umfüllung mit Schlauch, möglichst in CO2-Atmosphäre (z.B. Umleeren Primärgärtank –> Sekundärgärtank/ oder Abfüllung in Bierflaschen), geringer Kopfraum in Bierflaschen (möglichst füllen oder ausschäumen!), Kronkorken mit O2-Absorber (Info), tiefe Temperaturen bei Lagerung (< 6 °C). Siehe ausführliche LOB-Brewing-Anleitung hier. Top-Info zum LOB (Low Oxygen Brewing).
-
Biere möglichst frisch geniessen!

Abb. 26. Haupthopfenformen.
oben: frische Hopfendolden, links: getrocknete Hopfendolden, unten rechts: Hopfenpellets. Hopfenextrakt: cf. hier.

Abb. 27. Aufbewahrung von Hopfen.
Links: Pellets vakuumiert, rechts: Pellets vakuumiert in einer Lichtschutz-Packung Lagerung: im Kühlschrank, besser im Tiefkühler. Vorgehen: siehe hier.

Abb. 28. Flaschenabfüllung.
Die Bierabfüllung in Flaschen ist kritisch bezüglich Sauerstoffeintrag.
1. Einlaufvorgang: Bier sollte am Boden der Flasche langsam, möglichst nicht blubbernd einlaufen.
2. Volumen Kopfraum (zwischen den Pfeilen):
Der Kopfraum sollte möglichst klein sein. 2ml Luft im Kopfraum einer 500ml Flasche erhöhen den Sauerstoffgehalt bereits um mehr als 1mg/L. Eine maximale Füllmenge reduziert somit wesentlich den Rest-O2-Gehalt.
Das minimale Restvolumen in der abgebildeten 33 cL-Bierflasche Longneck mit einer Abfüllpistole Metall (Info) beträgt 18.5 mL.
3. CO2-Ausspülung: Ideal wäre die Ausspülung der Flasche beim Abfüllvorgang mit CO2 bzw. einem CO2/N2-Gasgemisch (vor dem Abfüllen).
4. Schaumbildung: Eine CO2-Schaumbildung Im Flaschenhals trägt ebenfalls zur Verdrängung des Luftsauerstoffs bei.

Abb. 25. Malz: ungeschrotet und geschrotet.
li: Ungeschrotetes Malz ist mit einer derben Frucht- und Samenschale gegen die O2-haltige Umwelt abgeschirmt. Die Inhaltsstoffe, insbes. die Enzyme sind vor Oxidationen gut geschützt.
re: Geschrotetes Malz hat eine viel grössere Oberfläche und keine Schutzhülle mehr. Unerwünschte oxidative Reaktionen laufen ab, Enzyme denaturieren.
Weiterführende Informationen und Anleitungen zum Thema Sauerstoff und Bier finden sich auf dieser
Website unter:
-
Theorie: Physikalisch-chemisches Braulabor > 3.10. Die Bestimmung des gelösten Sauerstoffgehalts
> 3.10.1. Unerwünschter Sauerstoff beim Brauprozess --> siehe hier
-
Theorie + Praxis: Informationen zur Auswirkung des "unerwünschten Sauerstoffs" auf Bier sowie detaillierte
Anleitung/Empfehlungen zur Vermeidung der Sauerstoffaufnahme während dem gesamten Brauprozess
("Low Oxygen Brewing") --> siehe pdf-Anleitung hier (S. 1 bis 3)
-
Praxis: Bestimmung des gelösten Sauerstoffgehaltes --> siehe pdf-Anleitung hier (S. 4 bis 8)
-
Theorie: Bierfehler Oxidation --> siehe hier
-
Praxis: Lagerung von Hopfen --> siehe hier.

LOD (Low Oxygen Drinking): vor zuviel Sauerstoffzutritt schützen!!
Literaturangaben
Bücher
- Annemüller, G., Manger, H.-J., Lietz, P. Die Hefe in der Brauerei. VLB, Berlin (2020)
- Annemüller, G., Manger, H.-J. Gärung und Reifung des Bieres. VLB Berlin (2013)
- Barth, R. The Chemistry of Beer. The Science in the Suds. John Wiley, Hoboken NY (2013)
- Berg, J.M., Tymoczko, J.L., Stryer, L. Biochemie. Spektrum Akademischer Verlag, Heidelberg (2007)
- Boulton, C., Quain, D. Brewing Yeast & Fermentation. Blackwell Science, Oxford UK (2006)
- Brücklmeier, J. Bier Brauen. Grundlagen, Rohstoffe, Brauprozesse. Eugen Ulmer, Stuttgart (2018)
- Kunze, W. Technologie. Brauer & Mälzer, VLB Berlin (2016)
- Palmer, J.J. Erfolgreich Bier brauen. Mobiwell Verlag, Immenstadt (2019)
- Sadava, D., Hillis, D.M., Heller, H.C., Berenbaum, M.R. Life. The Science of Biology. Sinauer/Freeman, Sunderland (2010)
- White, C., Zainasheff, J. Yeast. The Practical Guide to Beer Fermentation. Brewers Publication, Boulder (2010)
Internetquellen
- Aquilla T. Aeration & Oxygenation Wort. MoreBeer! [Quelle]
- Ebbinghaus, U., Briem, F. Was Bier kaputt macht. Frankfurter Allgemeine 22.02.2028 [Quelle]
- Fix, G.J. Controlling Beer Oxidation. Brewing Techniques [Quelle]
- O`Rourke, T. The role of oxygen in brewing [Quelle]
- Staudt, A. Oxidation und Bieralterung. Brau!magazin Winter 2014/15 [Quelle]
- Stika, J. Aerating Wort Technique. BYO [Quelle]
- The Modern Brewhouse. Methods of the modern brewhouse (LOB) [Quelle]
- ThermoFischer Scientific. Beer Brewing & Dissolved Oxygen [Quelle]
- Verbelen, P.J. et al. The influence of yeast oxygenation prior to brewery fermentation on yeast metabolism and the oxidative stress response. FEMS Yeast Res 9, 226-239 (2009)
- Wyeast. Oxygenation/ Aeration [Quelle]

MUG
Mikrobrauerei
seit 2012
Für Edge+Chrome+Firefox auf Notebook, Personal Computer und iPad optimiert, für Smartphones ungeeignet
06.02.2026